О.А. Коваленко, к. с.-г. н., зав. каф. рослинництва та садово-паркового господарства Миколіївського НАУ,
А.І. Ковбель, голова ФГ «Деметра +»
Для життя та розвиту рослині впродовж всього вегетаційного періоду необхідна певна кількість макро-, мезота мікроелементів, а для їх споживання – певні ґрунтовокліматичні умови: структура ґрунту, його температура та вологість, рН, температура та вологість повітря, кількість та інтенсивність сонячної радіації та інші.
Зміна клімату в сторону потепління та зменшення кількості атмосферних опадів, часті ґрунтові та повітряні посухи вводять рослини в стресовий стан. Ми не можемо змінити природні фактори, але маючи в розпорядженні низку агротехнічних та агрономічних прийомів, можемо вплинути на імунітет рослин, підвищити стресостійкість, посухостійкість, оптимізувати використання наявних елементів живлення та вологи для формування вегетативної маси та генеративних органів.
Мікроелементи є основою життя. Всі процеси синтезу та перетворення речовин здійснюються за допомогою ферментів, до складу яких входять мікроелементи. Вони відіграють значну роль у процесах синтезу білків, жирів, вуглеводів, а також входять до складу вітамінів, гормонів та інших біологічно активних речовин. Очевидним є той факт, що нема жодного більш чи менш важливого біохімічного процесу, жодної фізіологічної функції, які б могли проходити без участі того чи іншого мікроелемента.
Наявність та можливість засвоєння елементів живлення, особливо в критичні фази розвитку рослин, вплив окремих елементів на адаптацію рослин до стресових умов дає можливість, коригуючи вегетативним внесенням потрібних елементів живлення, впливати на структуру елементів урожайності рослин.
Одним із найважливіших мікроелементів для формування урожайності є бор. Він істотно впливає на вуглеводний і білковий обмін у рослинах, на утворення фітогормонів – ауксинів. Бор – генератор клітин – активізує поділ клітин та розвиток молодих тканин, підвищує еластичність стінок клітин, бере участь у синтезі нуклеїнових кислот ДНК і РНК, впливає на ріст кореневої системи, особливо молодих коренів, зумовлює активність ферментів, сприяє синтезу хлорофілу та асиміляції вуглекислого газу. За дефіциту бору порушується перехід вуглеводів і крохмалю з листків в інші органи рослини, гальмується процес фотосинтезу, розвиток кореневої системи, а також розвиток точки росту (зокрема, меристематичних клітин).
При позакореневому підживленні рослин мікродобривами та монохелатами важливим фактором їх ефективності є фази розвитку рослин на момент їх внесення, відповідно до цього – вплив на фізіологічні процеси, від яких залежать елементи урожайності. Бор безпосередньо впливає на ріст пилкових трубок та проростання пилку і його визрівання, що забезпечує запилення та запліднення квіток, збільшення кількості плодів і насіння. Коригуючи вміст бору в вегетуючих рослинах впродовж їх онтогенезу, можна впливати на їх продуктивність, якщо інші фактори, що є не менш значущими (макроелементне живлення, вологозабезпечення та ін.), також враховані. Бор майже не рухається з нижньої частини рослини до точки росту (не реутилізується), його дефіцит зумовлює відставання рослини в розвитку. За дефіциту бору пилок є стерильним і формування зернівки не відбувається. Стійкість рослин до стресових ситуацій (спека, посуха) значно знижується.
Позакореневе підживлення рослин мікроелементами оптимізує макроелементне живлення. При достатньому забезпеченні мікроелементами рослина створює більше вегетативної маси на 1520% (в сухій речовині), споживаючи на 20% менше вологи.
Живлення рослини відбувається шляхом поглинання кореневою системою елементів живлення з ґрунту. При дефіциті поживних речовин недостатньо розвивається коренева система, як наслідок, замала її асиміляційна поверхня, а значить, і площа живлення. Коли температура в зоні кореневої системи нижча за +14оС, рослина не здатна засвоїти в повному обсязі наявні в ґрунтовому розчині поживні елементи. Так, при температурі +14оС в зоні життєдіяльності кореневої системи рослина зможе засвоїти лише 20% від наявних у ґрунтовому розчині рухомих форм фосфору. В умовах посухи, коли ґрунтовий розчин практично відсутній, відбувається порушення макроелементного живлення: калій і фосфор рослиною не засвоюються, порушується метаболізм азоту, накопичується етилен, який призводить до старіння рослин, зумовлюючи незворотні процеси початку гибелі рослин.
Скоригувати макрота мікроелементне живлення можна шляхом позакореневого підживлення рослин відповідними комплексами мікроелементів та функціональними добривами для позакореневого підживлення. Зокрема, переліку цих вимог відповідає лінійка мікроелементних та функціональних добрив торгової марки Quantum, що випускає ТОВ НВК «Квадрат».
Препарат КВАНТУМФІТОФОС містить спеціальні формуляції фосфору і калію (композиція фосфітів (PO3) і фосфатів (PO4) та калію в суміші з органічними кислотами). Фосфати, надходячи в рослину, відразу включаються в обмін речовин. Таким чином, відбувається корекція фосфорного живлення в критичні фази розвитку рослин і при його нестачі.
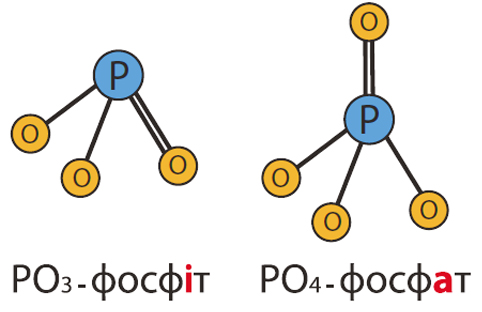
Фосфіти, маючи на один атом кисню менше, володіють унікальною мобільністю по ксилемі і флоемі, тому легко проникають в усі частини рослини, стимулюють розвиток кореневої системи, збільшуючи розміри та кількість кореневих волосків. Фосфіти дуже легко проникають через листкову поверхню рослин і дуже погано споживаються кореневою системою при наявності їх в ґрунтовому розчині (Lowatt and Mikkelsen, 2006).
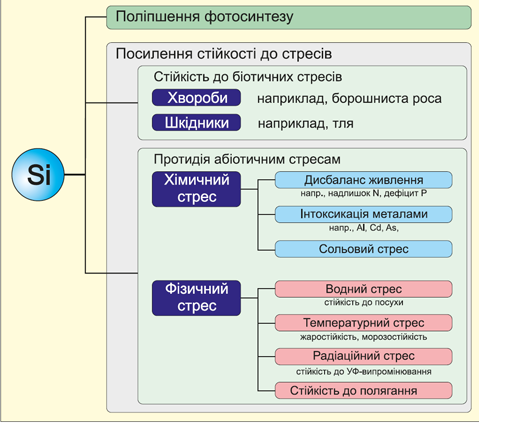
Позитивний вплив кремнію на ріст і розвиток рослин забезпечується підвищенням стійкості до біотичних та абіотичних стресів. Джерело: Trends Plant Sci. 2006 Aug; 11(8):392-7.
Фосфіт, що знаходиться в рослині, блокує збудників грибних захворювань (Phytophtora, Mildew, Pythium, Fusarium, Rhizoctonia), не допускаючи утворення спор (фунгіцидний ефект) і спричиняє імунну відповідь рослини виробленням фітоалексинів та інших вторинних метаболітів. Фосфіти, володіючи яскраво вираженим фунгіцидним ефектом, є джерелом фосфору пролонгованої дії. Під дією певних факторів фосфіт перетворюється на фосфат і є додатковим джерелом живлення (Lovatt 1990). При гострому дефіциті фосфору фосфіт, потрапляючи в рослину, відразу включається в метаболізм (Lowatt and Mikkelsen, 2006).
Калій є рухомим елементом і знаходиться в сольовій формі в клітинному сокові. Від його концентрації залежать рух води в листі рослин та інтенсивність дихання. Калій регулює процеси транспірації і дихання, контролюючи відкриття і закриття устячок, тому при достатній кількості калію підтримується тургор (вміст рідини в клітинах) і скорочуються втрати води (в’янення рослин) в умовах посухи та спекотної погоди. Калій сприяє росту судин ксилеми, покращуючи таким чином можливість транспорту цукрів і накопичення вуглеводів. Калій має велике значення для регулювання процесів транспірації дихання, він поліпшує стійкість рослин до стресу в умовах посухи і високих температур. Високоефективна формуляція «органічного» калію забезпечує ефективне і швидке поглинання через листкову поверхню. По поширеності кремній є другим (після кисню) елементом земної кори і ґрунту.
Незважаючи на те що кремній поглинається рослинами і грунтовими мікроорганізмамив у кількостях, що перевищують величину поглинання основних макроелементів (N, P, K), сьогодні його не відносять до числа первинних поживних речовин вищих рослин, хоч Ю.Лібіх у 1840 році поставив кремній четвертим елементом живлення після азоту, фосфору, калію. Тим не менше, позитивна роль кремнію в стимуляції росту і розвитку багатьох рослин загальновідома. При цьому позитивний ефект кремнію особливо помітний у рослин в стресових умовах. (Епштейн, 1994, 1999, 2009; Ma, 2004, 2006). Зазначимо деякі закономірності, які виділяють кремній з ряду інших елементів в життєдіяльності рослин. Майже всі рослини (за незначними винятками) можуть бути вирощені без кремнію в поживному середо вищі, навіть кремнієфільні рис і пшениця! Другою особливістю є те, що діапазон концентрацій кремнію в різних рослинах значно ширший, ніж у інших поживних елементів. Так, вміст кремнію коливається в діапазоні 0,110% від маси сухої речовини, в той час як, наприклад, для азоту цей діапазон становить 0,56%, для калію – 0,88%, фосфору – 0,150,5%. Тобто різниця концентрацій кремнію – 100 раз!, а других елементів – не більше 10 раз! (Епштейн, 1994).
Дослідження дозволили зробити висновок, що рослини можуть поглинати низькомолекулярні кремнієві кислоти та їх аніони не тільки через кореневу систему, а й через поверхню листків, якщо обприскувати їх кремнієвмісними водними розчинами. Важливо зазначити, що поглинання кремнію листками складає близько 3040%, тоді як через кореневу систему не перевищує 15% (Матиченков В.В., 2008). Кремній відкладається у вигляді прошарку товщиною 2,5 мкм в просторі безпосередньо під тонким (0,1 мкм) шаром кутикули, утворюючи подвійний кутикулярний кремнієвий шар на поверхні листків. Окрім того накопичення кремнію відбувається також в епідермісі та провідних тканинах стебла, листків і оболонки зерен. Однією із важливих функцій активних форм кремнію є стимуляція розвитку кореневої системи (Кудинова, 1975; Adatia, Besford, 1986).
Дослідження на злакових, цитрусових, овочевих культурах і кормових травах засвідчили, що при покращенні кремнієвого живлення рослин збільшується кількість вторинних і третинних корінців на 20100% і більше. Дефіцит кремнієвого живлення є одним із лімітуючих факторів розвитку кореневої системи рослин. Встановлено, що оптимізація кремнієвого живлення призводить до підвищення стійкості молекул хлорофілу (Matichenkov et al., 2004; Іванов та ін., 2005) і самої концентрації хлорофілу (Wang S.Y. та ін., 1998).
Особливу і дивовижну роль відіграє кремній у підвищенні стійкості рослин до стресів різноманітного походження (як біотичних, так і абіотичних). Дослідження та висновки передових світових вчених в останнє десятиліття висувають саме цю властивість кремнію на перше місце (Liang Y., 2007; E. Epstein, 2009; Heather A. Currie, 2007; Матиченков, 2008; та ін.)
В цій системі хімічного захисту беруть участь також речовини та первинні метаболіти, що беруть участь в основному обміні речовин рослини – фітогормони, органічні кислоти та ін. Яка ж роль кремнію в цій «хімічній війні»? Саме кремній бере безпосередню участь у біосинтезі захисних метаболітів і на сьогодні цьому є незаперечні докази (Матиченков, 2008).
В літературі висунуто кілька механізмів, що пояснюють підвищення посухостійкості рослин при оптимізації кремнієвого живлення. (Ahmad et al,, 1992; Aston & Jones, 1976; Ma & Takahashi, 2002). Один із механізмів пов’язаний з підвищенням водоутримувальної властивості ґрунтів при внесенні кремнієвмісних сполук. Іншим поясненням може служити пониження транспірації та зміна кута нахилу листків рослин, що забезпечує менший рівень випаровування вологи, які відбуваються завдяки підвищеній акумуляції кремнію рослинами. Проте такий визначний отриманий ефект не можна пояснити дією тільки цих механізмів. Очевидно, підвищення стійкості рослин до дефіциту вологи в проведеному експерименті пов’язане перш за все з прямою дією активних форм кремнію на рослини. Накопичувана рослиною монокремнієва кислота може полімеризуватися як у клітинному, так і в позаклітинному просторі рослинних тканин. Si має властивість накопичувати і зберігати воду в організмі рослини завдяки формуванню полікремнієвих кислот та їх гелів. Дослідження засвідчили, що один атом Si в таких гелях може утримувати до 146 молекул води. Із врахуванням «витрат» поглинутого рослиною кремнію на формування «скелета» і захисного шару в епідерміс було визначено, що близько 2030% наявного в організмі кремнію може брати участь у процесі підтримки внутрішнього резерву води (Воронков та ін., 1978; Ahmad et al., 1992).
Компанія «Квадрат» виробляє препарат Квантум-АкваСил, що містить 20% кремнію, 8-10% калію та 1% гумінових речовин.
2012 рік був надзвичайно посушливим і тому показовим при застосуванні даної схеми використання комплексних мікроелементних та функціональних добрив торгівельної марки Quantum на товарних та дослідних посівах, де досліджувався вплив варіантів допосівного обробітку зерна та вегетаційного внесення різних варіантів захисностимулюючих композицій, їх вплив на структуру елементів урожайності та врожайність у цілому.
Для прикладу: урожайність кукурудзи при допосівному обробітку зерна протруйником та препаратом Квантум-КУКУРУДЗА на контролі – 32,4 ц/ га (1), при вегетаційному підживленні препаратами Квантум-КУКУРУДЗА + Квантум-ХЕЛАТ ЦИНКУ + Квантум-БОР АКТИВ + «Квантум-ФІТОФОС» – 55,0 ц/ га (2), при додаванні в бакову суміш додатково препарату «Квантум-АкваСил» – 65,0 ц/га (3).
Аналізуючи отримані експериментальні дані, помічаємо суттєвий позитивний вплив у даних ґрунтовокліматичних умовах на елементи врожайності та врожайність в цілому, використання в вегетаційних підживленнях комплексних хелатних мікроелементних добрив, монохелатів та функціональних добрив. Застосування препарату Квантум-БОР АКТИВ суттєво вплинуло на озернення початків кукурудзи. Застосувавши препарат Квантум-ФІТОФОС, ми покращили баланс макроелементного живлення, провели профілактичне лікування та зміцнили імунітет рослин (2). В результаті цього рослини краще розвивалися, були стійкішими до стресових умов, зокрема, недостатньої вологозабезпечуваності та високої температури, що наочно видно на тепловому фотознімку, де температура оброблених рослин більш ніж на 2оС нижча, ніж на контролі (1).
Використання препарату Квантум-АкваСил сприяло збільшенню урожайності кукурудзи на 10 ц/га (3) завдяки підвищенню імунітету і оптимізації водного балансу рослини та зменшенню транспірації рослинної вологи, підвищенню стійкості рослин до УФ-опромінення. Рослини продовжували вегетацію на 20 днів довше, ніж на контролі, а температура вегетуючих рослин була на 4оС нижчою ніж у рослин, що не були оброблені.

1 2 3

1

2

3



